Inbreeding is a result of the mating of individuals which are related to one another by having one or more common ancestors. If the mated individuals are related, their offspring will to some extent be inbred.
The coefficient of inbreeding
The rate of inbreeding depends on the degree of relationship
Inbreeding also arises as a result of restricted population size
Full sib mating
Inbreeding depression
Heterosis or hybrid vigor
Inbred strains
References
The coefficient of inbreeding
The coefficient of inbreeding, is the probability that the two genes at any locus are identical by descent (Falconer 1981), i.e. that the two genes are copies of one of the genes carried by the common ancestor a few generations back. The coefficient of inbreeding, symbolised by F, is a property of an individual, but inbreeding profoundly effects the genetic composition of a population and in appropriate circumstances can lead to the formation of inbred strains in which all individuals are virtually genetically identical.
The rate of inbreeding depends on the degree of relationship
The closest relationship is that of an individual with itself, or self- fertilisation. However, the closest relationship that is usually possible with mammals is full brother x sister (known as full-sib) mating. Continuous mating of offspring to the younger parent (which prevents repeated backcrossing to the same individual, which would have different genetic consequences), or a single generation of parent x offspring mating is genetically equivalent to full-sib mating.
Other regular mating systems which lead to a high level of inbreeding include half-sib and cousin matings. Repeated backcrossing, say of a transgene or a new mutation, to an inbred strain increases homozygosity as rapidly as self-fertilisation.
Inbreeding also arises as a result of restricted population size
In a closed colony it eventually becomes impossible to avoid the mating of related individuals. Hence even “outbred” stocks maintained as a closed colony gradually become inbred at a rate which depends on the size of the colony. The rate of inbreeding with different sizes of colonies is shown graphically in Fig. 1. Mathematical explanations of the consequences of brother x sister mating and other regular systems of inbreeding are given elsewhere (Falconer 1981, Green 1981). Contrary to popular belief, avoiding brother x sister mating in a small closed, random-mated population may reduce the inbreeding of an individual but it does not reduce the over-all rate of inbreeding. This is because the inbreeding will be undone in a subsequent generation.
Inbreeding is always expressed relative to an arbitrary starting point at which the coefficient of inbreeding is assumed to be zero. Therefore, the magnitude of the effects of inbreeding any specific population will depend on the previous history of the stock, and the extent to which it has already been inbred.
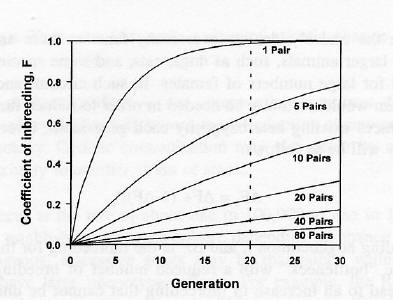
Fig. 1. Inbreeding as a result of restricted population size. A strain is regarded as an “inbred strain” when the coefficient of inbreeding, F, is greater than 0.986, i.e. after 20 generations of sib-mating.
Full sib mating
Full sib inbreeding of a genetically heterogeneous stock doubles the total genetic variation if all the sublines are kept. However, all the genetic variation will then be due to differences between sublines, with no genetic variation within sublines (Falconer 1981).
The phenotypic variation among sublines also increases. This is largely due to the “uncovering” of recessive genes and genetic drift in which alleles at a particular polymorphic locus become fixed in a homozygous state with plus or minus (with respect to the character) alleles being fixed largely by chance. The phenotypic variation among a set of inbred strains derived from an outbred stock is therefore substantially greater than the phenotypic variation within the starting population.
The converse of this is also true. If several inbred strains are mixed together, then the phenotypic variation in the combined population will be less than that of the individual inbred strains taken together.
The coefficient of inbreeding never quite reaches 100 per cent. Therefore, no strain is ever fully inbred. Moreover, the coefficient of inbreeding is calculated on the assumption that the reproductive performance of heterozygotes is equal to that of homozygotes, and that no mutation occurs. Both of these assumptions are incorrect, and lead to a slight overestimate of the actual level of inbreeding. On the other hand, it is assumed that the base population has a coefficient of inbreeding of zero. In practice, many inbred strains are derived from outbred stocks which may have been maintained as closed colonies with a restricted population size for many generations, as a result of which they may already be highly inbred.
Inbreeding depression is a decline in reproductive performance, ability to survive and other characteristics associated with fitness as a result of inbreeding. It occurs as a result of “uncovering” deleterious recessive genes by making them homozygous and is a consequence of the evolution of dominance of loci concerned with fitness characters. The direction of the change is towards the value of the more recessive alleles (Falconer 1981). Inbreeding depression does not occur for those characters where the heterozygote is intermediate between the two homozygotes.
The degree of inbreeding depression depends on the previous history of the stock. A stock which has been kept as a closed population for many generations will already be partly inbred; hence, full-sib mating may not result in much inbreeding depression.
Inbreeding depression varies substantiallyamong different lines. Anyone starting a new inbreeding project should do so on a sufficiently large scale to allow for extinction of a proportion of the lines during the first few generations. Once an inbred strain has been established, no further inbreeding depression should occur. Any decline in breeding performance will be due either to environmental influences (particularly disease) or in some cases to new deleterious mutations becoming fixed in the strain.
Heterosis or hybrid vigor
Heterosis or hybrid vigor is an increase in fitness and its components which is observed in F1 hybrids between two inbred strains. It is the opposite of inbreeding depression, and results from the “covering up” of mildly deleterious recessive genes present in a homozygous state in the parental inbred strains. As with inbreeding depression, it is only seen in those characteristics where there is a dominant/recessive mode of inheritance. The effects of heterosis on breeding performance can be spectacular. A comparison of the breeding performance of five inbred mouse strains and the 20 hybrids obtained by crossing them in all possible combinations found that the inbred strains produced an average of 10.4 and the hybrids 19.1 young in the first two litters, and the best of the inbreds was worse than the worst of the hybrids (Festing 1976).
Inbred strains
The decision as to whether a strain is sufficiently inbred for any particular research project is largely arbitrary. The Committee on Standardized Genetic Nomenclature for Mice decided in 1952 that 20 generations of full-sib mating (or its genetic equivalent), at which time F= 98.6 per cent, is the minimum level of inbreeding required before a strain of mice can be designated as an inbred strain.
However, the coefficient of inbreeding never quite reaches 100 per cent so no strain is ever fully inbred. Moreover, the coefficient of inbreeding is calculated on the assumption that the reproductive performance of heterozygotes is equal to that of homozygotes, and that no mutation occurs. Both of these assumptions are incorrect, and lead to a slight overestimate of the actual level of inbreeding. But it is also assumed that the base population has a coefficient of inbreeding of zero. Many inbred strains are derived from outbred stocks which have been maintained as closed colonies with a restricted population size for many generations, as a result of which they may already be highly inbred.
There are an estimated 30,000 loci in the mouse, and about 35% (i.e. about 10,500) of these are normally polymorphic. After 20 generations of sib-mating, about 273 loci are expected still to remain unfixed within a strain, provided the base population really is genetically variable. Only after 40 generations of sib-mating is the number reduced to that of mutation load of about 6-12 loci (i.e. the genes still varying in the strain due to new mutations) (Bailey 1982). Thus, in some ways, it might have been better to require 40 generations of brother x sister mating before a strain can be designated as “inbredâ€. The chance of a locus of interest to a particular investigator still segregating is low.
The 20 generations of sib-mating standard has been accepted for most other laboratory species, although in the case of larger species such as the chicken, rabbit and pig, lower levels of inbreeding may be sufficient for some research purposes.
Inbreeding is defined in terms of the probability of heterozygosity at a locus. However, all inbred strains used in biomedical research should also be isogenic, i.e. all individuals within an inbred strain should be genetically identical (apart from residual segregation due to the impossibility of achieving fully inbred strains). In fact it is isogenicity rather than homozygosity that is the most useful property of inbred strains, and the two are distinct properties which should not be confused. Isogenicity is achieved by ensuring that all individuals trace back to a common ancestral full-sib breeding pair in the twentieth or a subsequent generation. All parallel substrains should be eliminated. F1 hybrids, i.e. the first-generation cross between two inbred strains, are isogenic but not homozygous. It is unfortunate that the terms inbred and `outbred’ which describe breeding methods rather than a genetic property of a group of animals, have become so widely accepted. The terms “isogenic” and “non-isogenic” are more useful in describing the genetic characteristics of these two types of laboratory animals.
References
Bailey DW (1982), How pure are inbred strains of mice?, Immunology Today 3: 210-214
Falconer DS. Introduction to quantitative genetics, 2nd. edition. 340pp. London, New York: Longmans., 1981.
Festing MFW. Effects of marginal malnutrition on the breeding performance of inbred and F1 hybrid mice-a diallel study. In: Antikatzides T, ed. The Laboratory Animal in the Study of Reproduction. Stuttgart: Gustav Fischer, 1976: 99-114.
Green EL. Genetics and probability in animal breeding experiments. 271pp. London and Basingstoke: The Macmillan Press Ltd., 1981.